




Moving Behavior of Adult Brown Planthoppers, Nilaparvata lugens
-
摘要:
目的 研究褐飞虱的移动行为规律,为褐飞虱行为调控防治提供新思路和理论支持。 方法 以室内续代饲养的稻飞虱品系为研究对象,采用自制的移动计数装置,对长翅和短翅两个性别褐飞虱成虫的移动行为规律进行了观察研究。 结果 4个虫态成虫的移动时间分布、移动量和移动昼夜比例等均存在明显差异。雄性成虫的移动频率显著高于雌性,其中长翅雄性成虫的移动总量最高,平均为1 290次;短翅雌性成虫最低,为480.7次。雄性成虫夜间移动量高于白天,而雌性特别是短翅型雌性成虫夜间移动量仅占0.36。 结论 褐飞虱成虫的移动行为具有一定规律,雌、雄成虫移动行为差异明显。根据4类成虫的移动行为差异和特点,通过人为方法干扰或抑制其移动行为,可能会影响到褐飞虱的迁飞和求偶活动,从而降低种群数量达到防治的目的。 Abstract:Objective Moving behavior of brown planthoppers, Nilaparvata lugens, one of the worst pests in rice fields, were studied. Method The moving behavior of adult macropterous as well as brachypterous male and female brown planthoppers were observed with a motion sensor and an automatic counter in an enclosed chamber. Result The 4 categories of brown planthoppers displayed significantly different patterns of motions. The males moved significantly more frequently than the females. The macropterous males were most active at 1290.0±11.93 times totally during the observation, while the brachypterous females at the minimum of 480.7±19.36.The males tended to move more frequently at night than during daytime; whereas, the females, under day light, particularly the brachypterous females which devoted only 36% of the activity at night. Conclusion There appeared repeat patterns for the different categories of brown planthoppers in moving around that could be of value for the pest control. -
Key words:
- behavioral pattern /
- wing type /
- sex difference /
- pest behavioral regulation
-
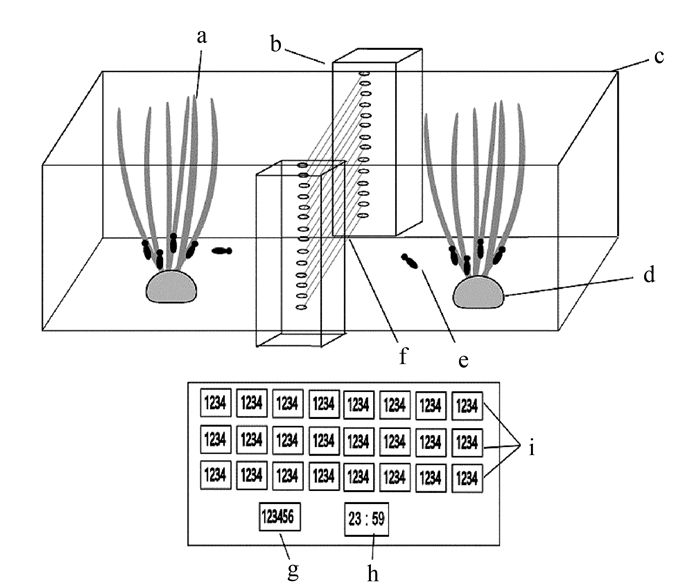
图 1 褐飞虱移动计数装置结构
注:a 为水稻幼苗;b 为红外光栅感应装置;c 为石英玻璃罩;d 为湿润棉花;e 为待测成虫;f 为红外光栅;g 为总量数字显示管;h 为计时数字显示管;i 为分时计数数字显示管。
Figure 1. Counting device for insect moving behavior
Note: a-rice seedlings; b-infrared grating sensor; c-quartz glass cover; d-wet cotton ball; e-tested insects; f-infrared rays; g-total number display; h-time display; and, I-hourly number display.

图 2 长翅雄性褐飞虱成虫各时段移动率
注:A为1、2日,B为3、4日,C为5、6日。图中阴影区域为夜间(无光照)时段;移动率=各时段移动量/当日移动总量。图 3~5同。
Figure 2. Hourly moving frequency of adult macropterous N. lugens males
Note:A.1, 2 d, B.3, 4 d, C.5, 6 d.Note: Shadow areas represent activities taken place in artificial night (without light); moving rate=moving frequency in an hour/total times of moving in a day.The same as Fig. 3-5.

图 3 长翅雌性褐飞虱成虫各时段移动率
Figure 3. Hourly moving frequency of adult macropterous N. lugens females
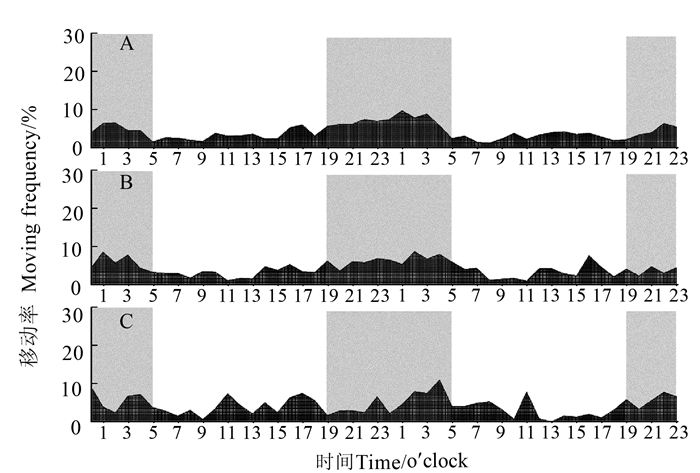
图 4 短翅雄性褐飞虱成虫各时段移动率
Figure 4. Hourly moving frequency of adult brachypterous N. lugens males
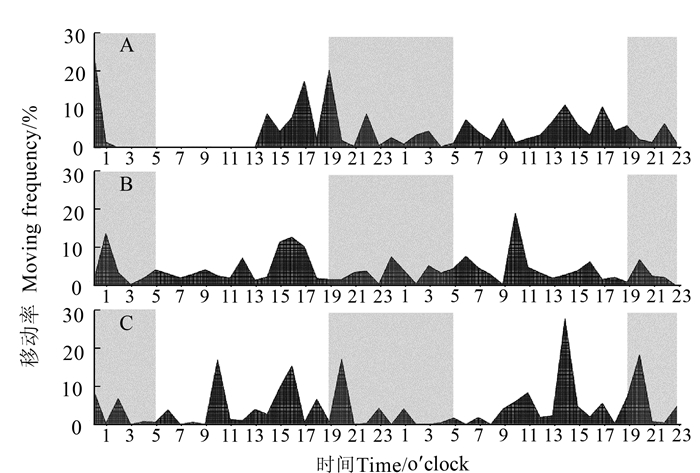
图 5 短翅雌性褐飞虱成虫各时段移动率
Figure 5. Hourly moving frequency of adult brachypterous N. lugens females
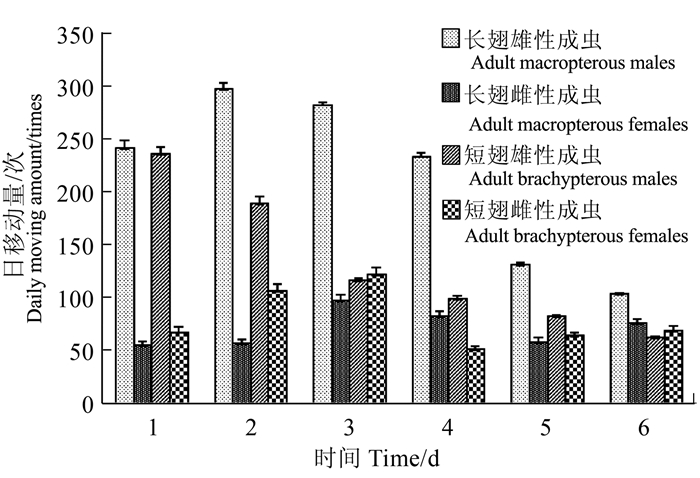
图 6 褐飞虱成虫单日移动量比较
Figure 6. Daily moving behaviors of adult N. lugens in different categories

图 7 褐飞虱昼夜分布比例比较
注:夜间移动比例=每日夜间移动量/日移动总量。
Figure 7. Ratios of day/night moving counts of adult N. lugens in different categories
Note: Nighttime moving rate = moving frequency at night/total moving counts on a same day.
表 1 褐飞虱成虫日移动差异比较
Table 1. Daily moving behaviors of adult N. lugens in different categories
变量
Source of VariationF值
F-ValueP值
P-Value组内变异 Within subject variation 观察时期 Observing time 5.097 0.004 观察时期*翅型 Observing time*Wing type 0.990 0.394 观察时期*性别 Observing time*Sex 3.842 0.014 观察时期*翅型*性别 Observing time*Wing type*Sex 1.115 0.037 组间变异 Between subject variation 翅型 Wing type 1.720 0.226 性别 Sex 11.649 0.009 翅型*性别 Wing type*Sex 2.646 0.142 注:重复测量方差分析,经球形检验P<0.05,采用Huynh-Feldt方法进行校正。
Note: Repetitive measurement and analysis of variance (RMANOVA), as the Mauchly's Test of Sphericity showed that P<0.05, a Huynh-Feldtcorrection method was used. 下载: 导出CSV
下载: 导出CSV
表 2 褐飞虱成虫移动总量差异比较
Table 2. Total moving counts of adult N. lugens in different categories
虫态
Insect state白天(光照)移动总量
Total moving events in daytime夜间(无光照)移动总量
Total moving events in darkness移动总量
Total moving events夜间移动比例
Moving ratio in darkness长翅雄性成虫 Adult macropterous males 521.7±2.26 a 768.3±10.82a 1290.0±11.93a 0.59±0.030 长翅雌性成虫 Adult macropterous females 213.3±12.18 c 213.3±10.92c 426.7±23.03c 0.51±0.045 短翅雄性成虫 Adult brachypterous males 366.3±8.46ab 420.3±3.08b 786.7±11.21b 0.55±0.044 短翅雌性成虫 Adult brachypterous females 300.3±10.75 bc 180.3±8.97c 480.7±19.36c 0.36±0.065 注:同一列标记字母不同者为差异显著(LSD多重比较,P<0.05)。*夜间移动比例=每日夜间移动量/日移动总量。
Note: Different letters within the same column represent significant differences, LSD, P<0.05.*The moving ratio in nighttime = the number of moving events in the nighttime of the day/the whole day amount of moving events.
下载: 导出CSV
-
[1] 李汝泽, 丁锦华, 胡国文.褐飞虱及其种群管理[M].上海:复旦大学出版社, 1996.LI R Z, DING J H, HU G W. Brown Planthopper and Its Population Management[M]. Shanghai:Fudan University Press, 1996.(in Chinese) [2] 程遐年, 吴进才, 马飞, 等.褐飞虱的研究与防治[M].北京:中国农业出版社, 2003.CHEN X N, WU J C, MA F, et al. Research and Control of Brown Planthopper[M]. Beijing:China Agriculture Press, 2003. (in Chinese) [3] 朱绍先, 邬楚中, 杜景祐.褐飞虱及其防治[M].上海:上海科学技术出版社, 1984.ZHU S X, WU C Z, DU J Y. Brown Planthopper and Its Control[M]. Shanghai:Shanghai science and technology press, 1984.(in Chinese) [4] 朱述钧.稻飞虱研究中几个热点问题的研究进展(综述)[J].安徽农业大学学报, 2006, 33:343-346. doi: 10.3969/j.issn.1672-352X.2006.03.013ZHU S J. Recent Advances in Some Important Research Areas About Planthoppers[J]. Journal of Anhui Agricultural University, 2006, 33:343-346.(in Chinese) doi: 10.3969/j.issn.1672-352X.2006.03.013 [5] 刘凤沂, 李惠陵, 邱建友, 等.惠州地区褐飞虱对几种药剂的抗药性监测[J].昆虫知识, 2010, 47:991-993. doi: 10.7679/j.issn.2095-1353.2010.182LIU F Y, LI H L, QIU J Y, et al. Monitoring of resistance to several insecticides in brown planthopper(Nilaparvata lugens) in Huizhou[J]. Chinese Bulletin of Entomology, 2010, 47:991-993.(in Chinese) doi: 10.7679/j.issn.2095-1353.2010.182 [6] 邱良妙, 林仁魁, 吴玮, 等. 6种杀虫剂对水稻褐飞虱田间种群的毒力及控制作用[J].福建农业学报, 2011, 26(3):424-431. doi: 10.3969/j.issn.1008-0384.2011.03.019QIU L M, LIN R K, WU W, et al. Toxicity and efficacy of six insecticides on field population ofNilaparvata lugens[J]. Fujian Journal of Agricultural Sciences, 2011, 26(3):424-431.(in Chinese) doi: 10.3969/j.issn.1008-0384.2011.03.019 [7] 凌炎, 黄凤宽, 龙丽萍, 等.中国和越南褐飞虱抗药性研究[J].应用昆虫学报, 2011, 48(5):1374-1380. http://d.old.wanfangdata.com.cn/Periodical/kczs201105031LIN Y, HUANG F K, LONG L P, et al. Studies on the pesticide resistant of Nilaparvata lugens(Stål) in China and Vietnam[J]. Chinese Journal of Applied Entomology, 2011, 48(5):1374-1380.(in Chinese) http://d.old.wanfangdata.com.cn/Periodical/kczs201105031 [8] 朱道弘.昆虫翅型分化的调控及翅多型性的进化[J].昆虫知识, 2009, 46(1):11-16. doi: 10.3969/j.issn.0452-8255.2009.01.003ZHU D H. Regulatory mechanism and evolution of insect wing polymorphism[J]. Chinese Bulletin of Entomology, 2009, 46(1):11-16.(in Chinese) doi: 10.3969/j.issn.0452-8255.2009.01.003 [9] 解再宏, 苏品, 廖晓兰.褐飞虱翅型分化影响因素及机制研究综述[J].江西农业学报, 2009, 21(10):95-99. http://d.old.wanfangdata.com.cn/Periodical/jxnyxb200910030XIE Z H, SU P, LIAO X L. Research on wing dimorphism of brown planthopper[J].Acta Agriculturae Jiangxi, 2009, 21(10):95-99.(in Chinese) http://d.old.wanfangdata.com.cn/Periodical/jxnyxb200910030 [10] 周崇高, 冯娅琳, 陆潮峰, 等.稻飞虱翅型分化的研究进展[J].湖北农业科学, 2014, 53(17):3985-3990. doi: 10.3969/j.issn.0439-8114.2014.17.001ZHOU C G, FENG Y L, LU C F, et al. Advances in wing dimorphism of rice planthoppers[J]. Hubei Agricultural Sciences, 2014, 53(17):3985-3990.(in Chinese) doi: 10.3969/j.issn.0439-8114.2014.17.001 [11] 张志涛, 陈伦裕.三种飞虱、叶蝉鸣声的采集和分析[J].科学通报, 1987, 32(20):1583-1583. http://www.cnki.com.cn/Article/CJFDTotal-KXTB198720016.htmZHANG Z T, CHEN L Y. Collection and analysis of the song of three planthoppers and leafhoppers[J]. Chinese Science Bulletin, 1987, 32(20):1583-1583.(in Chinese) http://www.cnki.com.cn/Article/CJFDTotal-KXTB198720016.htm [12] 姚青, 赖凤香, 傅强, 等.褐飞虱求偶鸣声中有效声段及其特征[J].昆虫学报, 2004, 47(4):479-484. doi: 10.3321/j.issn:0454-6296.2004.04.012YAO Q, LA F X, FU Q, et al. Effective sections and sensitive characters of courtship signals in the rice brown planthopper, Nilaparvata lugens (Stål)[J]. Acta Entomologica Sinica, 2004, 47(4):479-484.(in Chinese) doi: 10.3321/j.issn:0454-6296.2004.04.012 [13] 程遐年, 陈若篪, 习学, 等.稻褐飞虱迁飞规律的研究[J].昆虫学报, 1979, 22(1):1-21. http://d.old.wanfangdata.com.cn/Periodical/stxb201408015CHEN X N, CHEN R C, XI X, et al. Studies on the migrations of brown planthopper Nilaparvata lugens Stål[J].Acta Entomologica Sinica, 1979, 22(1):1-21.(in Chinese) http://d.old.wanfangdata.com.cn/Periodical/stxb201408015 [14] KISIMOTO R. Meteorologica conditions inducing long-distance immigration of the brown planthopper, Nilaparvata lugens (Stål)[J]. Chinese JEnt, 1984, 4(2):39-48. [15] 陈若篪, 程遐年.褐飞虱起飞行为与自身生物学节律、环境因素同步关系的初步研究[J].南京农业大学学报, 1980, 3(2):42-49.CHEN R C, CHEN X N. The take-off behavior of brown planthopper (Nilaparvata lugens Stål) and its synchronous relations to the biological rhythm and environmental factors[J].Journal of Nanjing Agricultural University, 1980, 3(2):42-49.(in Chinese) [16] OH R J. Repeated copulation in the brown planthopper, Nilaparvata lugens Stál (Homoptera; Delphacidae)[J]. Ecol Entomol, 1979(4):345-353. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=10.1111/j.1365-2311.1979.tb00594.x [17] NOVOTNY V. Adaptive significance of wing dimorphism in males of Nilaparvata lugens[J]. Entomol Exp Appl, 1995, 76:233-239. doi: 10.1111/j.1570-7458.1995.tb01967.x [18] 张志涛, 殷柏涛, 陈伦裕.褐稻虱发声机理模型和雌虫鸣声的人工模拟[J].中国水稻科学, 1991, 5(1):29-36. doi: 10.3321/j.issn:1001-7216.1991.01.006ZHANG Z T, YIN B T, CHEN L Y. The model of signal production and the simulation of female signals in rice brown planthopper, Nilaparvata lugens(Stål)(Homootera:Delphacidae)[J]. Chinese Journal of Rice Science, 1991, 5(1):29-36.(in Chinese) doi: 10.3321/j.issn:1001-7216.1991.01.006 -
 点击查看大图
点击查看大图
计量
- 文章访问数: 940
- HTML全文浏览量: 225
- PDF下载量: 19
- 被引次数: 0
