




Chemistry and In Vitro Immunological Activity of Polysaccharides from Sparassis latifolia
-
摘要:
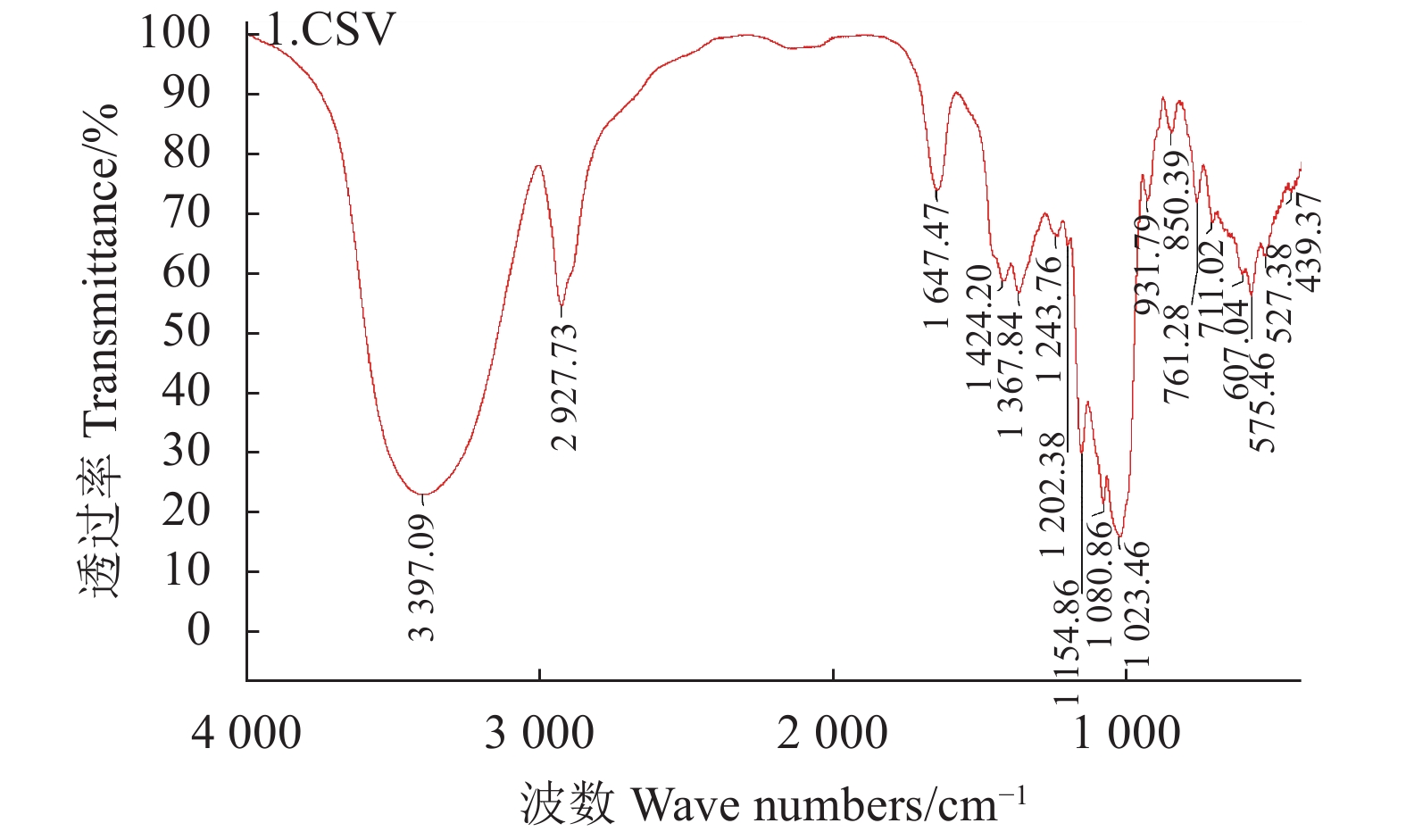
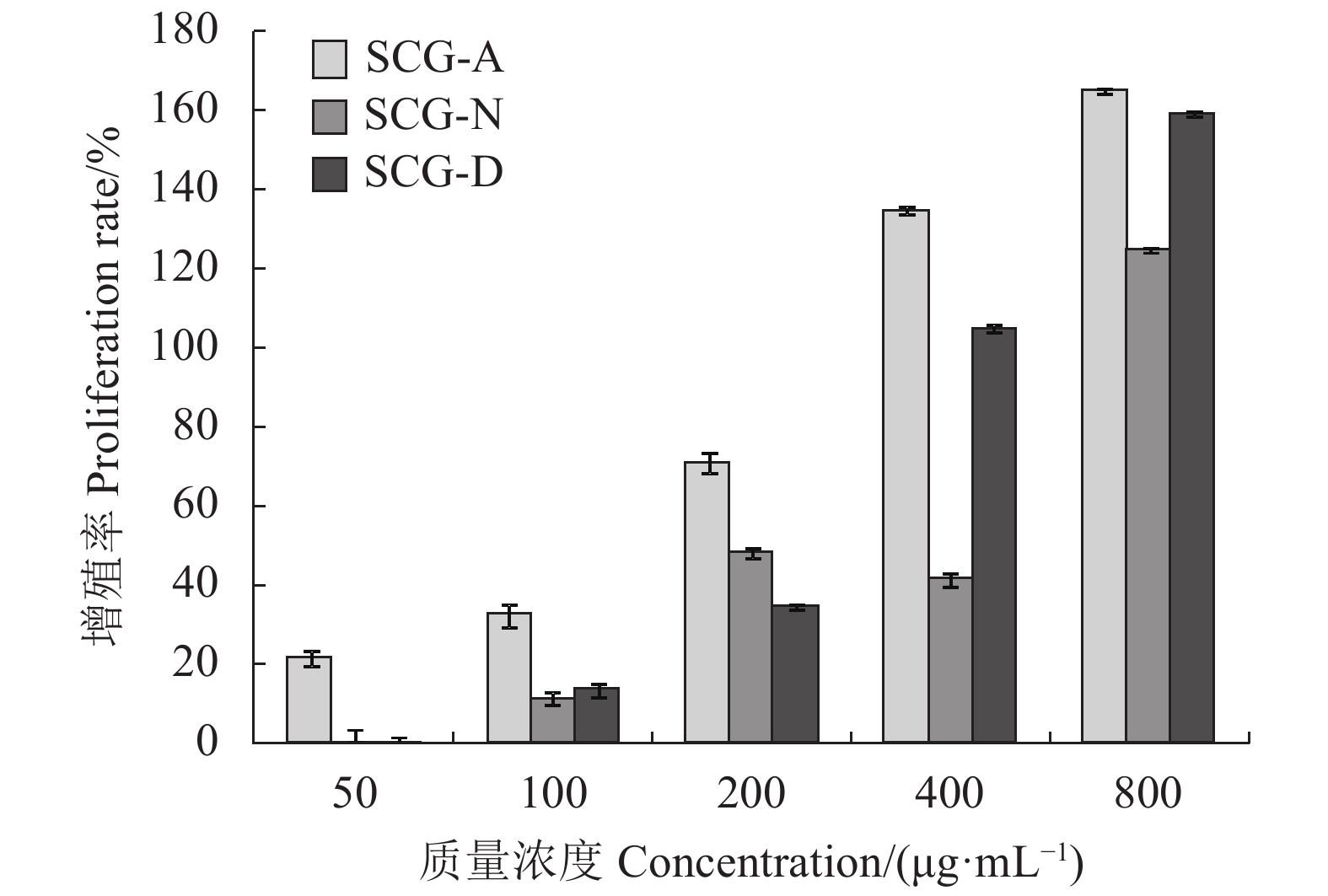
目的 分析绣球菌水溶性多糖的结构与免疫活性。 方法 采用热水提取法从广叶绣球菌Sparassis latifolia的真空冷冻干燥品中提取得到绣球菌水溶性多糖SCG-D,然后通过DEAE Sepharose Fast Flow离子交换层析对其进行分离,获得SCG-N和SCG-A两个多糖组分,并对SCG-A组分进行HPSEC、单糖组成分析、红外光谱、1H-NMR和13C-NMR核磁共振的结构分析。通过大鼠脾淋巴细胞体外刺激活性测试比较了SCG-D及其组分SCG-A、SCG-N的体外免疫活性差异。 结果 绣球菌水溶性多糖SCG-D具有促进鼠脾淋巴细胞增殖的活性,其酸性多糖组分SCG-A促进作用最强,其100 μg·mL−1 72 h处理组的增殖率为32.7%,而SCG-N多糖组分的活性较SCG-A低,其100 μg·mL−1 72 h处理组增殖率为11.66%。结构分析结果表明SCG-A的重均分子量为4.30×105 Da,其主链结构为α-1,4-D-葡聚糖,主要由葡萄糖和半乳糖构成,并且还含有一定量的1-6分支结构。 结论 绣球菌冻干品的水溶性多糖及其组分具有促进大鼠脾淋巴细胞体外增殖的免疫活性,其中活性最强的酸性多糖组分SCG-A是一种具有1-6分支结构的α-1,4-葡聚糖。 Abstract:Objective Chemical structure and in vitro immunological activity of the water soluble polysaccharides from Sparassis latifolia were studied . Method The lyophilized powder of S. latifolia fruiting bodies was extracted by hot water to obtain polysaccharides SCG-D. From the extract, the SCG-N and SCG-A fractions were isolated by the DEAE-Sepharose Fast Flow ion exchange chromatography. The structure of SCG-A was analyzed using HPSEC, monosaccharide composition analysis, FT-IR, and NMR. The in vitro immunological activities of the extract and the fractions were determined by a test on rat spleen lymphocytes. Result SCG-D was found active in promoting the lymphocyte proliferation. SCG-A exhibited the strongest activity with a proliferation rate of 32.7% at the concentration of 100 μg/mL in 72h, while SCG-N activity was lower at 11.66%. The molecular weight of SCG-A was 4.30×105Da with an α-1,4-D-glucan as its main chain consisting primarily of glucose and galactose with some 1-6-branches. Conclusion The water-soluble polysaccharides and its fractions obtained from the lyophilized S. latifolia powder showed varying in vitro immune activities of promoting the proliferation of rat spleen lymphocytes. The acidic SCG-A had a main chain structure of α-1,4-glucan with 1-6-branches and showed the greatest immunological activity among the 3 polysaccharide materials. -
Key words:
- Sparassis latifolia /
- polysaccharide /
- spleen lymphocyte /
- immunity
-
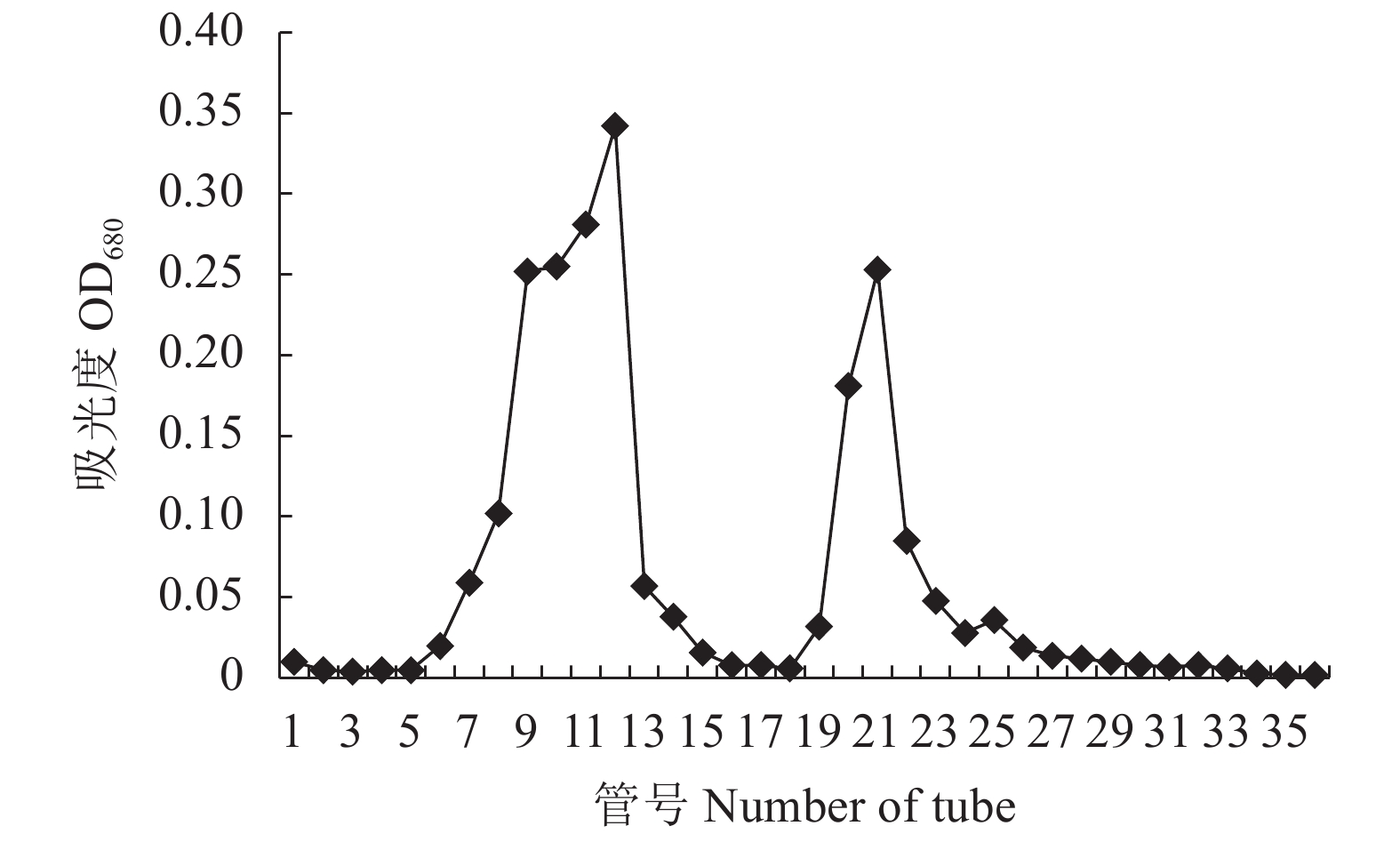
图 1 绣球菌SCG-D多糖DEAE Sepharose Fast Flow的洗脱曲线
Figure 1. Elution curve of SCG-D on DEAE Sepharose Fast Flow resin
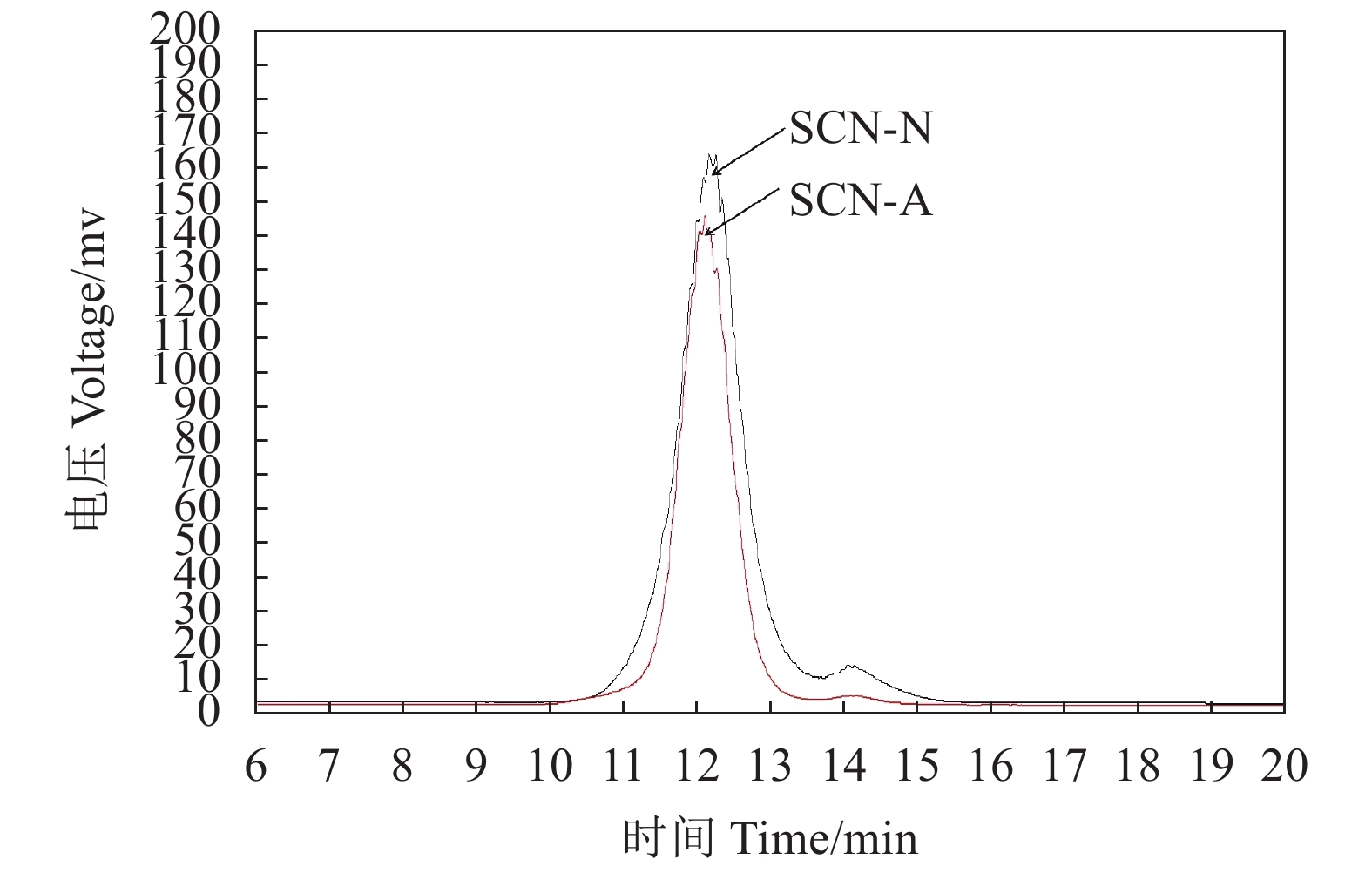
图 2 SCG-A和SCG-N的HPSEC凝胶色谱图
Figure 2. HPSEC gel chromatography of SCG-A and SCG-N fractions

图 3 标样与SCG-A水解样品PMP衍生物的色谱图
注:1:甘露糖;2:鼠李糖;3:葡萄糖醛酸;4:半乳糖醛酸;5:葡萄糖;6:半乳糖;7:木糖。
Figure 3. Chromatography of PMP derivatives of monosaccharide standards and SCG-A hydrolysate
Note: 1: Man; 2: Rha; 3: GlcU; 4: GalU; 5: Glu; 6: Gal; and, 7: Xyl.
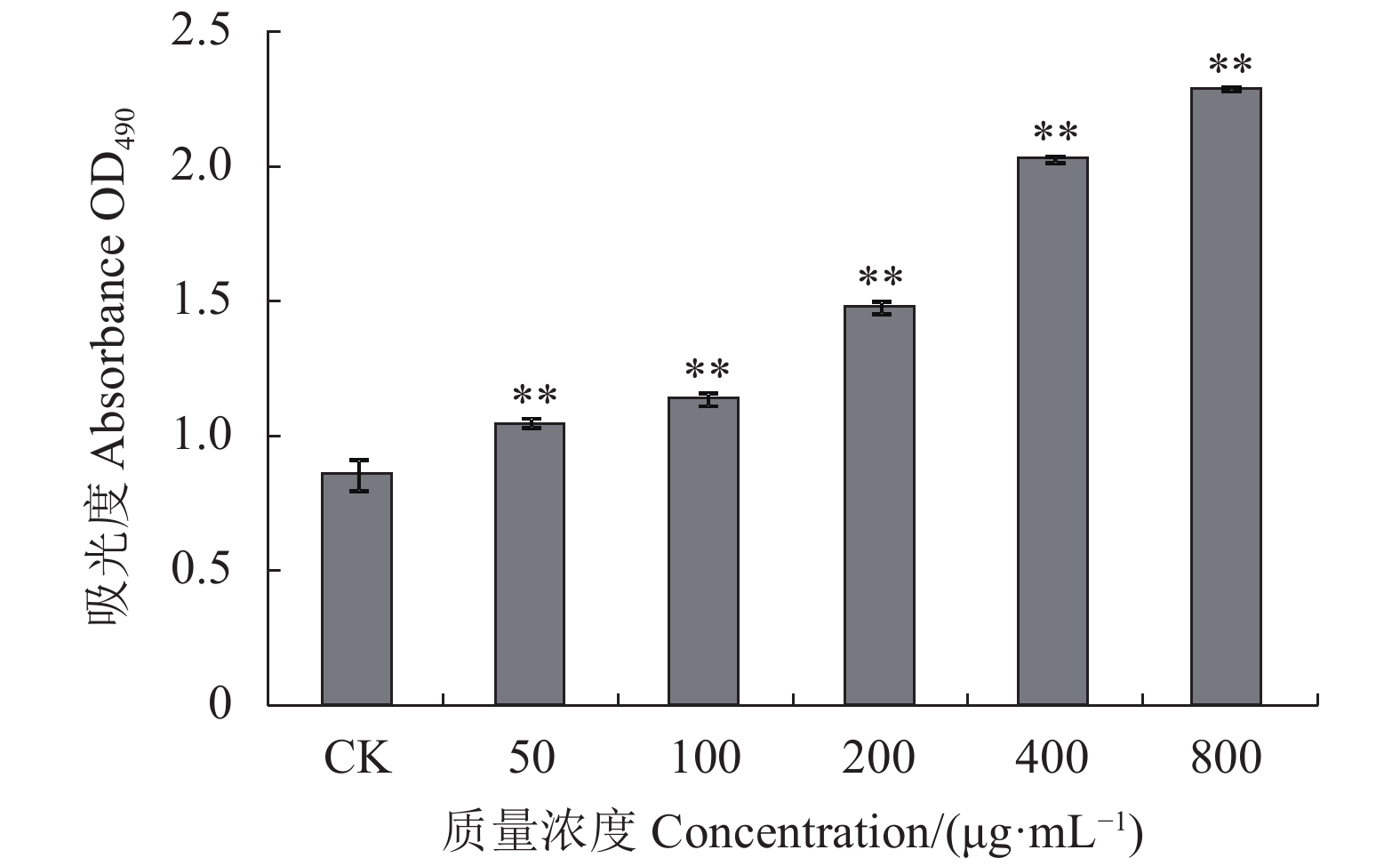
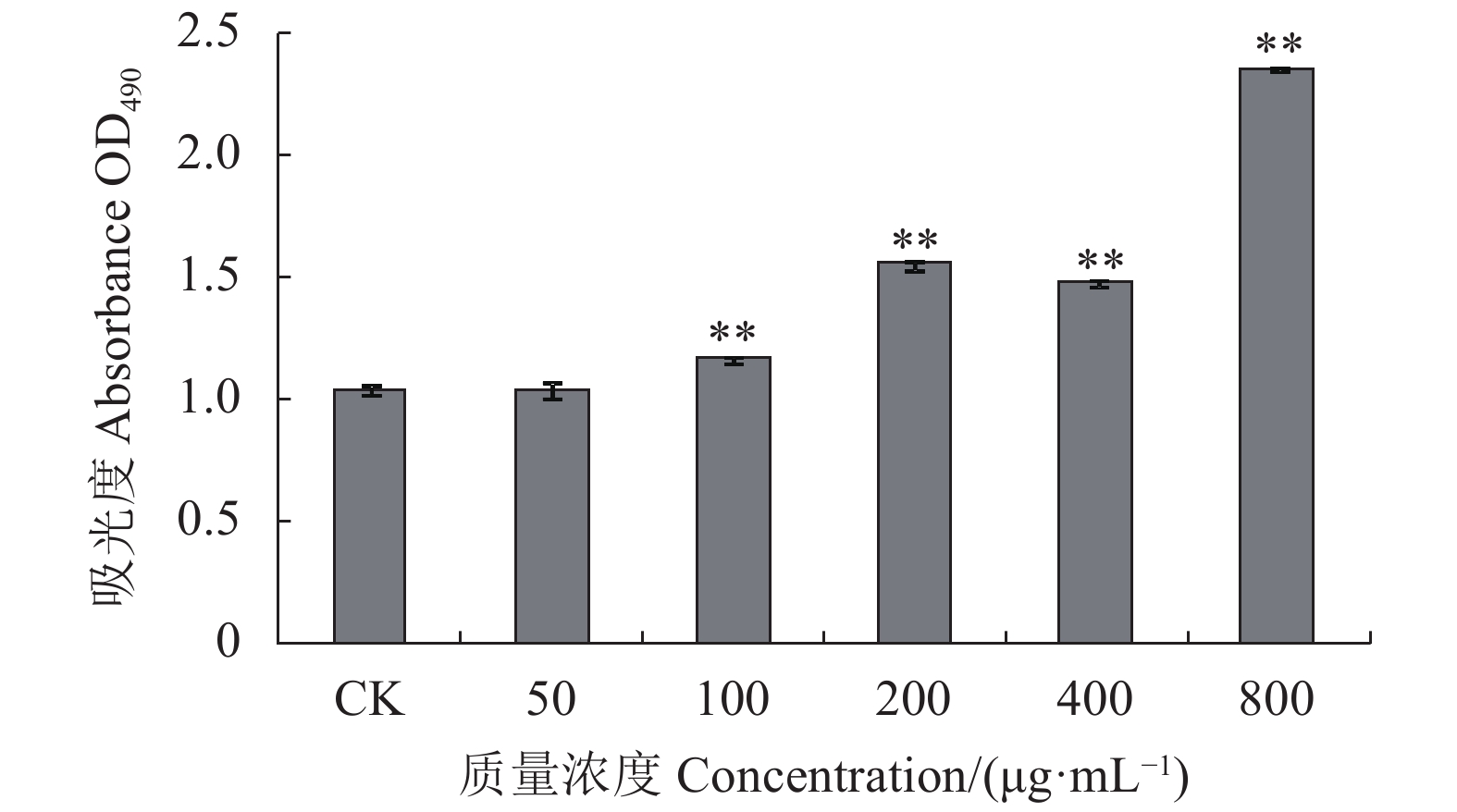
图 8 绣球菌多糖SCG-N对大鼠脾淋巴细胞增殖的影响
Figure 8. Effect of SCG-N on in vitro proliferation of rat spleen lymphocytes
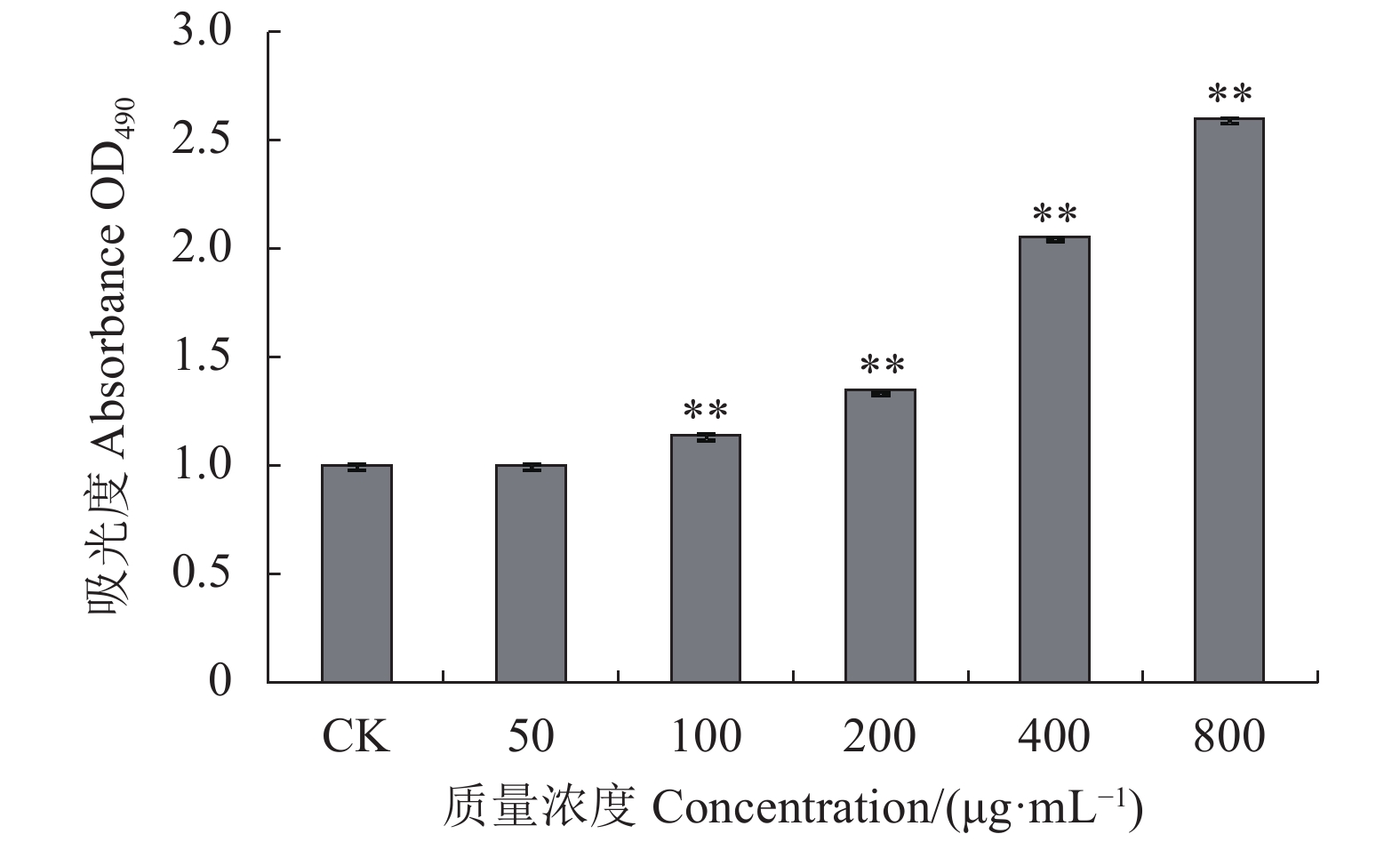
图 9 绣球菌多糖SCG-D对大鼠脾淋巴细胞增殖的影响
Figure 9. Effect of SCG-D on in vitro proliferation of rat spleen lymphocytes
-
[1] KIMURA T. Natural products and biological activity of the pharmacologically active cauliflower mushroom Sparassis crispa [J]. BioMed Research Intermational, 2013, 8(3): 501−508. [2] CHANDRASEKARAN G, OH D S, SHIN H J. Properties and potential applications of the culinary-medicinal cauliflower mushroom, Sparassis crispa (Wulf.: Fr.) (Aphyllophoromycetideae): a review [J]. International Journal of Medicinal Mushrooms, 2011, 13(2): 177−183. doi: 10.1615/IntJMedMushr.v13.i2.100 [3] KIM H S, KIM J Y, RYU H S, et al. Induction of dendritic cell maturation by β-glucan isolated from Sparassis crispa [J]. International Immunopharmacology, 2010, 10(10): 1284−1294. doi: 10.1016/j.intimp.2010.07.012 [4] KIM H H, LEE S, SINGH T S, et al. Sparassis crispa Suppresses mast cell-mediated allergic inflammation: Role of calcium, mitogen-activated protein kinase and nuclear factor-κB [J]. Intermational Journal of Molecular Medicine, 2012, 30(2): 344−350. doi: 10.3892/ijmm.2012.1000 [5] 白晨, 宋文荣, 杨剑飞, 等. 绣球菌子实体干燥条件与多糖保留率相关性研究 [J]. 食品科学, 2012, 33(20):119−122.BAI C, SONG W R, YANG J F, et al. Effect of Drying Methods on Retention Rate of Polysaccharides in Sparassia crispa Fruit Bodies [J]. Food Science, 2012, 33(20): 119−122.(in Chinese) [6] 朱美静, 童群义. 猴头多糖脱蛋白方法的研究 [J]. 河南工业大学学报(自然科学版), 2005, 26(4):25−27. doi: 10.3969/j.issn.1673-2383.2005.04.007ZHU M J, TONG Q Y. Study on Removal of Proten from Hericium erinaceus Polysaccharides [J]. Journal of Henan University of Technology (Natural Science Edition), 2005, 26(4): 25−27.(in Chinese) doi: 10.3969/j.issn.1673-2383.2005.04.007 [7] 金丽琴, 薛胜霞, 吕建新, 等. 牛膝多糖衍生物对小鼠脾淋巴细胞增殖及诱生IL-2和TNF-α的影响 [J]. 中国生化药物杂志, 2008, 29(5):312−314.JIN L Q, XUE S X, LU J X, et al. Effect of derivatives of Achyranthes bidentata polysaccharides on lymphocyte proliferation and induction of IL-2 and TNF- [J]. Chinese Journal of Biochemical Pharmaceutics, 2008, 29(5): 312−314.(in Chinese) [8] JAMAS S, EASSON D, STROFF GRO, et al. Method for producing soluble glucan: United States Patent, 5633369[P]. 1997. [9] TZIANABOS A O. Polysaccharide immunomodulators as therapeutic agents: structural aspects and biologic function [J]. Clinical Microbiology Reviews, 2000, 13(4): 523−533. doi: 10.1128/CMR.13.4.523 [10] 张迪, 王宏雨, 林衍铨. 不同干制方法对广叶绣球菌多糖提取率和分子量分布的影响 [J]. 中国食用菌, 2017, 36(3):54−56.ZHANG D, WANG H Y, LIN Y Q. Effects of Different Drying Methods on Extraction Rate and Molecular Weight Distribution of Polysaccharides from Sparassis latifolia [J]. Edible Fungi of China, 2017, 36(3): 54−56.(in Chinese) [11] 钱正明, 李文庆, 孙敏甜, 等. 冬虫夏草化学成分分析 [J]. 菌物学报, 2016, 35(4):476−490.QIAN Z M, LI W Q, SUN M T, et al. Analysis of chemical compounds in Chinese cordyceps [J]. Mycosystema, 2016, 35(4): 476−490.(in Chinese) [12] 马晓丽, 孟磊, 孙莲, 等. HPLC分析大蒜多糖中的单糖 [J]. 中国现代应用药学杂志, 2009, 26(7):585−587.MA X L, MENG L, SUN L, et al. Determination of Monosaccharide Compositions and Contents in Polysaccharide of Garlic by HPLC [J]. Chinese Journal of Modern Applied Pharmacy, 2009, 26(7): 585−587.(in Chinese) [13] 李平厉. 硫酸化可德兰多糖的制备、体外免疫调节、抗乙肝病毒感染活性及体内用作乙肝疫苗[D]. 济南: 山东大学, 2014.LI P L. Studies on the preparation, in vitro immunoregulatory andanti-hepatitis B virus infection activities and invivo action as a hepatitis B vaccine adjuvant of curdlan sulfate[D]. Jinan: Shandong University, 2014. (in Chinese) [14] 陈晓兰, 杨海峰, 瞿静雯, 等. 不同桑叶提取物对小鼠脾脏淋巴细胞增殖能力的影响 [J]. 中国畜牧兽医, 2017, 44(12):3598−3604.CHEN X L, YANG H F, QU J W, et al. Effect of Different Mulberry Leaf Extracts on Spleen Lymphocyte Proliferation in Mice [J]. China Animal Husbandry & Veterinary Medicine, 2017, 44(12): 3598−3604.(in Chinese) [15] RUI TADA, TOSHIE HARADA, NORIKO NAGI-MIUR, et al. NMR characterization of the structure of a b-(1, 3)-D-glucan isolate from cultured fruit bodies of Sparassis crispa [J]. Carbohydrate Research, 2007, 342: 2611−2618. doi: 10.1016/j.carres.2007.08.016 -
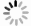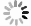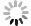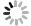
 下载:
下载:

 点击查看大图
点击查看大图
计量
- 文章访问数: 1223
- HTML全文浏览量: 531
- PDF下载量: 13
- 被引次数: 0
